Recent Changes in Hammerhead
Taxonomy
Glenn A. Fines wrote: "I just have a quick question for someone who may have more literary resources available than myself. I am working on a presentation on Hammerhead sharks. All of the literature I have (1993 and earlier) classify this group as Family Sphyrnidae with two genera Sphyrna and Eusphyra. However, it was mentioned to me that this may have been changed recently and the family Sphyrnidae was reverted to a single genus Sphyrna with a subgenus Eusphyra within the family Carcharhinidae. If anyone has an up to date classification, or knows where I can find one, of the hammerheads I would greatly appreciate it."
A lot has happened in hammerhead systematics and taxonomy in recent years. As yet, the nomenclatural dust has not settled, but I will try to bring you (and anyone else who's interested) up to speed as best I can.
The eight extant species of hammerhead shark (Sphyrna couardi was synonymised under S. lewini by McEachran and Serret in 1986) are among the most easily recognised of sharks. To a greater or lesser extent, all hammerheads are characterized by unique lateral expansions of the orbital and nasal regions of the chondrocranium and associated tissues, forming a distinctive double-bladed 'cephalofoil'.
Other than this unusual headgear, the hammerheads are very similar in overall form to the whaler sharks of the family Carcharhinidae (as established by Jordan & Evermann in 1896). Most modern workers (such as Bigelow & Schroeder 1948, Gilbert 1967, Compagno 1973, 1979, 1988, Bass, D'Aubray, & Kistnassamy 1975, Nelson 1976, 1984, Cadenat & Blanche 1981, among others) therefore regarded the hammerheads as a distinct family, the Sphyrnidae sensu Gill 1872, closely allied with and almost certainly derived from the Carcharhinidae. Compagno (1979, 1988) noted that the hammerheads are morphologically most similar to the carcharhinid Scoliodon laticaudus (the spadenose shark) and hypothesised that the hammerhead sharks may share a relatively recent common ancestor with that genus. The earliest hammerheads in the fossil record of which I am aware are undescribed teeth in the collection of David Ward from middle Eocene deposits of Tunisia but the Scoliodon/Loxodon/Rhizoprionodon to Sphyrna transition is extremely difficult to determine precisely, as the teeth of these genera are very similar (D. Ward, pers comm.).
It had been assumed (Compagno 1979, 1988) that the hammerheads represent a monophyletic group (including all the descendants of a common ancestor) and that the cephalofoil evolved gradually, starting from a non-hammerheaded carcharhinoid ancestor most probably similar to Scoliodon in overall form. According to this common-sense scenario, the basic 'hammer' design evolved only once in sharks, the modestly-cephalofoiled bonnethead shark (Spyrna tiburo) was the earliest of the hammerheads to diverge and the boomerang-nosed winghead shark (Eusphyra blochii I'll get into the matter of genera and subgenera momentarily) the most recent.
|
|
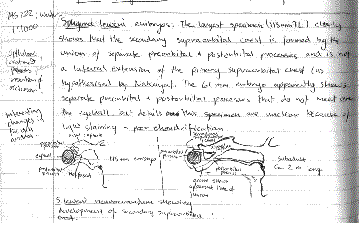
Based on my own dissections of the heads of embryo, juvenile, and adult hammerheads of several species, I seemed to see evidence that the hammer did indeed evolve gradually from smaller to larger. The sequence of hammer evolution from none to modest to extreme seemed to make perfect sense and to be fully supported by morphological and developmental evidence. But it now seems that our collective common sense was simply wrong. Using mtDNA sequences, Martin (1993) provided powerful evidence that the hammerheads are indeed a monophyletic group but that the sequence of divergence was precisely opposite from that which had long been assumed: the 'extremely' hammer-headed winghead shark was the earliest of the group to diverge and the comparatively ridge-headed bonnethead the most recent. Further, Martin's analysis suggested that the relative width of the cephalofoil in various species of hammerheads (expressed as a percentage of body length) does not show a clear trend in the in-between species of hammerheads. This pattern suggests that the hammerhead cephalofoil appeared rather suddenly, fully realized, and was only later modified in various ways in response to differing selective pressures.
A simplified version of Martin's (1993) cladogram:
- - - - - - - - - - - - - -> [Representative Carcharhinids]
\- - - - - - - - - - - - -> Eusphyra blochii Winghead
Shark
\- - - - - - - - - - - -> Sphyrna mokarran Great Hammerhead
\- - - - - - - - - - -> Sphyrna lewini
Scalloped Hammerhead
\- - - - - - - - - -> Sphyrna corona Scalloped
Bonnethead
\ /- - -> Sphyrna media
Scoophead Shark
\ - - - - |
\ \- - -> Sphyrna tudes Smalleye Hammerhead
\ - - -
- - -> Sphyrna tiburo Bonnethead Shark
In a popular article originally published in Sea Frontiers (it has appeared in several forms since then), I tried to answer the question of why hammerhead sharks have such bizarre heads that is, what 'function' the hammer might serve these sharks. As I noted, there are basically two schools of thought on the matter: 1) hydrodynamic benefit versus 2) sensory enhancement. Evidence for the former school includes: hammerheads are among the most negatively buoyant of sharks (Baldridge 1970, 1972); a relatively constant ratio of anterior planing surfaces (there seems to be an inverse relationship between the size of the hammer and the pectoral fins, resulting in a fairly constant total surface area of these planing surfaces), the wing-like cross-section of the hammer (obvious on inspection), possibly acting as a canard; and the apparent autonomic control hammerheads seem to have over the trailing edge of the cephalofoil (only recently published, see Nakaya 1995), noting that other sharks often seem to avoid hammerheads in competitive contexts, possibly because of their enhanced maneuverability. Evidence for the latter school includes: early arrival at olfactory attractants (possibly due to wide spacing of the nares, allowing hammerheads to sample a wider portion of the water column) and observations by myself and others of hammerheads using their hammer in mine-sweeper fashion, apparently to locate prey buried in the sediment using the electrosensitive organs (ampullae of Lorenzini) distributed over much of the undersurface of the head.
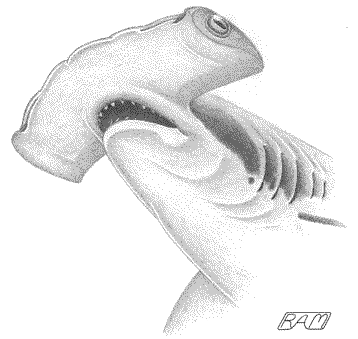 As a specialist in the behavioral ecology of elasmobranchs, I was (and still
am) somewhat biased in favor of the second school of thought that the
hammerhead 'hammer' was originally more a matter of sensory enhancement than
hydrodynamic benefit. Using the example of the 'extremely' hammer-headed winghead shark, I inferred that the 'cephalofoil' in this species seemed
downright unwieldy, apparently developed to a point beyond any conceivable
hydrodynamic benefit, yet the nares (which, unlike other species of hammerhead,
are closer to the midline of the hammer than the tips) and ampullae (though not
as widely spread over the undersurface of the hammer as in other species) are
more widely separated than in most non-hammerheaded sharks. I further noted that
the hammerhead cephalofoil is a complex structure that probably serves more than
one function. In conclusion, I therefore ranked the functions of the
hammerhead cephalofoil (arranged from most to least significant) as: 1)
increasing electrosensitive acuity, 2) increasing scent-tracking efficiency, 3)
increasing maneuverability, and 4) increasing lift. Although I had gotten the
sequence of hammerhead divergence bass-ackward, Martin's (1993) findings do seem
to support my bias that the hammer originally evolved as a sensory structure,
only later taking advantage of hydrodynamic advantages accruing from lateral
expansion of the head. The way I now interpret the sequence of hammerhead
divergence is as follows: The hammer appeared relatively suddenly (possibly the
result of relatively minor changes in epistatic genes or perhaps but less
likely transporons changing the rate or pattern of developmental events during
epigenesis), resulting in a hammer similar to that we see in the modern winghead
shark (which can be up to 44% of the standard length, with the nares located
closer to the midline of the hammer than the tips and the ampullae of Lorenzini
clustered mostly near the center of the hammer); in later 'models', the hammer was
decreased in size, the nares migrated to the tips of the hammer and the pattern
of ampullae become more complex and widespread over the undersurface of the
hammer (see Gilbert 1967, p 70, for an illustration of this). I therefore submit
that the hammerhead 'hammer', once it appeared, became fixed under conditions
of enhanced prey-locating ability (which would be expected to result in enhanced
reproductive fitness) and was later modified to take advantage of hydrodynamic
benefits as a secondary consideration. Therefore, in the development of the
hammerhead cephalofoil, sensory enhancement was initially more important than
hydrodynamic benefit.
As a specialist in the behavioral ecology of elasmobranchs, I was (and still
am) somewhat biased in favor of the second school of thought that the
hammerhead 'hammer' was originally more a matter of sensory enhancement than
hydrodynamic benefit. Using the example of the 'extremely' hammer-headed winghead shark, I inferred that the 'cephalofoil' in this species seemed
downright unwieldy, apparently developed to a point beyond any conceivable
hydrodynamic benefit, yet the nares (which, unlike other species of hammerhead,
are closer to the midline of the hammer than the tips) and ampullae (though not
as widely spread over the undersurface of the hammer as in other species) are
more widely separated than in most non-hammerheaded sharks. I further noted that
the hammerhead cephalofoil is a complex structure that probably serves more than
one function. In conclusion, I therefore ranked the functions of the
hammerhead cephalofoil (arranged from most to least significant) as: 1)
increasing electrosensitive acuity, 2) increasing scent-tracking efficiency, 3)
increasing maneuverability, and 4) increasing lift. Although I had gotten the
sequence of hammerhead divergence bass-ackward, Martin's (1993) findings do seem
to support my bias that the hammer originally evolved as a sensory structure,
only later taking advantage of hydrodynamic advantages accruing from lateral
expansion of the head. The way I now interpret the sequence of hammerhead
divergence is as follows: The hammer appeared relatively suddenly (possibly the
result of relatively minor changes in epistatic genes or perhaps but less
likely transporons changing the rate or pattern of developmental events during
epigenesis), resulting in a hammer similar to that we see in the modern winghead
shark (which can be up to 44% of the standard length, with the nares located
closer to the midline of the hammer than the tips and the ampullae of Lorenzini
clustered mostly near the center of the hammer); in later 'models', the hammer was
decreased in size, the nares migrated to the tips of the hammer and the pattern
of ampullae become more complex and widespread over the undersurface of the
hammer (see Gilbert 1967, p 70, for an illustration of this). I therefore submit
that the hammerhead 'hammer', once it appeared, became fixed under conditions
of enhanced prey-locating ability (which would be expected to result in enhanced
reproductive fitness) and was later modified to take advantage of hydrodynamic
benefits as a secondary consideration. Therefore, in the development of the
hammerhead cephalofoil, sensory enhancement was initially more important than
hydrodynamic benefit.
Gilbert (1967) placed all hammerheads in the genus Sphyrna and recognized three subgenera within the genus, 1) Eusphyra for S. blochii, 2) Platysqualus for S. corona, S. media, S. tiburo, and S. tudes, and 3) Sphyrna for S. couardi (which is now a junior synonym of S. lewini), S. lewini, S. mokarran, and S. zygaena. Compagno (1988) erected Mesozygaena as a new subgenus for S. tudes, S. corona, and S. media, and retained S. tiburo in the subgenus Platysqualus and S. lewini, S. mokarran, and S. zygaena in the subgenus Sphyrna. Comapgno (1979, 1988) ranked Eusphyra Gill 1862 as a genus, based on the following characters: lateral blades of cephalofoil very narrow and wing-like; nostrils tremendously enlarged, width 0.75 to 0.88 of internarial width and nearly twice mouth width; prenarial nodules present. Dingerkus (1986) objected to the elevation of Eusphyra to generic status, on the grounds that it implies that Sphyrna is paraphyletic (does not include all the descendants of a common ancestor as in the case of 'reptiles', as usually defined, excludes birds and mammals, yet all three classes share a common ancestor). This argument is inherently circular (essentially complaining "Compagno can't split hammerheads into two genera, because then we won't have one genus of hammerheads"). It is worth noting in passing that the species rank is the only 'natural' taxon that can be clearly defined among sexually-reproducing organisms; all other taxa are arbitrary and may be regarded as most useful when they best serve their twin purposes (a basis for generalization in comparative studies and an information storage system see Mayr 1981 for a discussion of this). As I understand the implications of the cladogram presented in Martin (1993), since the winghead shark does not occur among the other hammerheads, assigning it to a separate genus (in this case, Eusphyra) does NOT render the genus Sphyrna paraphyletic. The real question, then, is: Are the differences noted by Compagno (1979, 1988) sufficiently great to warrant the winghead shark a genus distinct from other hammerheads? Nelson (1994) regarded Eusphyra as a subgenus; Shirai (1996) retained it as a full-fledged genus. As all biological taxa above the species rank have no 'reality' in the natural world, this is largely a subjective decision.

Until recently, the Scalloped Hammerhead (Sphyrna lewini) was thought to be distinct from the Whitefin Hammerhead (Sphyrna couardi). Despite its name, the Whitefin Hammerhead does not have white or white-tipped fins. On the basis of broadly overlapping morphometric characters, this species was synonymized under Sphyrna lewini by John McEachran and Bernard Seret in 1987 (Cybium, 11[1]: 39-46)
Photo © David Fleetham david@davidfleetham.com; used with the gracious permission of the photographer.
I have no claims to being a systematist or taxonomist; like many shark enthusiasts, I regard these matters occasionally interesting but mostly they are just plain necessary important scientific disciplines that enable us to be certain we are all discussing the same species or group within the same conceptual (usually evolutionary) framework. I am by inclination a 'lumper', preferring to simplify matters by grouping things into as few categories as serves my purposes. While I feel that Compagno sometimes divides groups of elasmobranchs too finely (in his 1984 FAO catalogue, Sharks of the World, 9 out of 30 [30%] families and 49 out of 98 (50%) genera are monotypic, represented by only a single species; given the extremely low genetic variability and tremendous genetic sluggishness of sharks in general, combined with their rather conservative structure, this in my admittedly inexpert opinion seems somewhat excessive), but Compagno's work is by all accounts very good and his conclusions have been borne out so often that I am very reluctant to dismiss his ideas without VERY compelling evidence to the contrary. To my mind, placing the winghead shark in its own genus, distinct from the other hammerheads, serves to reflect its unique cephalic features and early evolutionary divergence from the common ancestor of the group as indicated by Martin's (1993) mtDNA data. So, pending further evidence to the contrary, I'm prepared to go along with Compagno on this point for the time being. You may choose not to; that is your right and privilege as a free-thinking adult.
And finally, the familial status of the hammerhead sharks appears to be no longer warranted. Naylor (1992) examined 17 presumed loci of structural proteins in 37 species of carcharhinoid shark. Despite only limited overlap between the genetic markers used and including many species not examined by Lavery (1992), both Naylor and Lavery's results fully support Compagno (1979, 1988)'s contention that the hammerhead sharks form a monophyletic group, closely allied with the genus Rhizoprionodon (sharpnose sharks). Unfortunately, neither Naylor nor Lavery tested the spadenose shark which Compagno (1979, 1988) hypothesized is the sister taxon to the hammerheads to determine its phylogenetic position among the other carcharhinoids tested. But Naylor's (1992) genetic sequence data also revealed something rather surprising: the family Carcharhinidae (as presently defined and constituted) is not monophyletic UNLESS the hammerheads are included within that family. To put this more bluntly: despite its many unique features, the Sphyrnidae is not a valid family. Surprising as this finding may be, Compagno (1988) explored this very possibility, proposing an alternate phylogenetic classification to allow for inclusion of the hammerheads within the family Carcharhinidae. Naylor (1992) advocated accepting Compagno's alternate hypothesis, pending further data. Nelson (1994) and Shirai (1996) have adopted this taxonomic arrangement. Martin (1993) further strengthens the argument in favor of accepting Compagnos (1988) classificatory scheme; this approach is followed here with the exception of placing S. media and S. tudes in the same subgenus as S. tiburo, as suggested by Martin's cladogram.
A Provisional Cladistic Classification of Hammerhead Sharks within the Family Carcharhinidae (Modified after Compagno 1988)
| Family Carcharhinidae | ||||||||
| Subfamily Galeocerdinae | ||||||||
| Genus | Galeocerdo - TIGER SHARK | |||||||
| Subfamily Carcharhininae | ||||||||
| Supertribe Sphyrnini | ||||||||
| Tribe Scoliodontini | ||||||||
| Genus | Scoliodon - SPADENOSE SHARK | |||||||
| Tribe Sphyrnini | ||||||||
| Genus | Eusphyra | |||||||
| Species | blochii - WINGHEAD SHARK | |||||||
| Genus | Sphyrna | |||||||
| Subgenus Sphyrna | ||||||||
| Species | lewini - SCALLOPED HAMMERHEAD | |||||||
| mokarran - GREAT HAMMERHEAD | ||||||||
| zygaena - SMOOTH HAMMERHEAD | ||||||||
| Subgenus Mesozygaena | ||||||||
| Species | corona - SCALLOPED BONNETHEAD | |||||||
| Subgenus Platysqualus | ||||||||
| Species | media - SCOOPHEAD SHARK | |||||||
| tudes - SMALLEYE HAMMERHEAD | ||||||||
| tiburo - BONNETHEAD SHARK | ||||||||
| Subspecies tiburo (Atlantic form) | ||||||||
| Subspecies vespertina (Pacific form) | ||||||||
| Supertribe Carcharhinini | ||||||||
| Tribe Rhizoprionodontini | ||||||||
| Genus | Loxodon* - SLITEYE SHARK | |||||||
| Rhizoprionodon - SHARPNOSE SHARKS | ||||||||
| Tribe Carcharhinini | ||||||||
| Subtribe Isogomphodontini | ||||||||
| Genus | Isogomphodon - DAGGERNOSE SHARK | |||||||
| Subtribe Triaenodontini | ||||||||
| Genus | Triaenodon - WHITETIP REEF SHARK | |||||||
| Subtribe Carcharhinini | ||||||||
| Genus | Carcharhinus** - REQUIEM SHARKS | |||||||
| Glyphis - RIVER SHARKS | ||||||||
| Lamiopsis - BROADFIN SHARK | ||||||||
| Nasolamia - WHITENOSE SHARK | ||||||||
| Negaprion - LEMON SHARKS | ||||||||
| Prionace - BLUE SHARK | ||||||||
| NOTES | * |
Several of Naylor's (1992) analyses suggest a close affinity between the little-known Loxodon macrorhinus and (of all things) the Reef Whitetip Shark, Triaenodon obesus, which is attractive because the former appears to be a rather base-level carcharhinid and a close evolutionary affinity with Triaenodon may help explain the latter's many intriguing triakid-like characters. The phylogenetic position of Triaenodon has been the subject of much recent taxonomic fiddling; Randall, Allen & Steene (1990) and Myers (1991) placed it in the family Hemigaleidae, which also seems to fit the genetic data fairly well IF the molecular systematic trees are collapsed via semi-strict consensus and/or character weighting. Further study of the matter is indicated. [Up to Loxodon] |
| ** | Based on independent cladistic analyses by Compagno (1988) and Naylor (1992), the validity of this genus is now rather doubtful, and the genus Carcharhinus as it now stands will almost certainly require being split into two or more genera. The cladograms of these workers agree in many respects, the 'Carcharhinus' sharks falling fairly readily into two broad groups that correspond reasonably well to Springer's (1950, 1951) scheme of dividing these sharks into smooth-backed Carcharhinus and ridge-backed Eulamia. Springer's (1950, 1951) scheme may therefore be a good place to start untangling these commercially important but tough to tell-apart sharks. [Up to Carcharhinus] |
I hope this helps.
Cheers,
R. Aidan Martin
Originally posted to SHARK-L Feb 24, 1998.
UPDATE: Since this was originally posted, I have developed a novel phylogenetic hypothesis that redefines Carcharhinidae in a way that favors retention of Sphyrnidae as a valid family and seems fully consistent with all data available to me. This hypothesis is presently being tested as part of a collaborative morphological/molecular systematic project which I am not at liberty to discuss at this time.
